

Most people assume paleontologists have the fossil record sorted out by now. Centuries of digging, mountains of data, DNA analysis, electron microscopy – surely we’ve figured out where everything belongs. But scattered across museum drawers and research journals are specimens that have survived every test modern science can throw at them and still refuse to give a straight answer. Not gaps waiting to be filled. Actual enigmas that get stranger the closer experts look.
Some of these fossils have been studied for over a hundred years. Some were reconstructed completely backward. One was assembled from parts scientists thought were three different animals. A few may represent entire branches of life that rose, dominated their world, and vanished without leaving a single living descendant – or even a recognizable relative. The 15 entries below are the ones keeping paleontologists up at night, and the deeper you go into this list, the weirder it gets.
#15 – The Tully Monster’s Stubborn Identity Crisis

The Tully Monster has been argued over since a fossil collector named Francis Tully pulled it out of an Illinois coal mine in 1958. Tullimonstrum gregarium sports a long proboscis ending in a claw, stalked eyes mounted on a rigid bar, and a soft segmented body that matches nothing alive or extinct with any confidence. For decades it sat in a cabinet as a curiosity. Then a 2016 study declared it a vertebrate – specifically, a jawless fish – based on what researchers interpreted as gill structures and a notochord. The paleontology world briefly celebrated having solved it.
The celebration didn’t last. In 2023, a team led by Tomoyuki Mikami at the University of Tokyo published results that directly challenged the vertebrate classification. Using 3D laser scanning on over 150 specimens, they reexamined the fossil’s internal structures – and concluded the Tully Monster lacked definitive vertebrate features. Instead, they suggested the Tully Monster may have been an invertebrate chordate – animals like lancelets or tunicates, which share a distant relationship with vertebrates but do not possess a backbone. Most unsettling of all, both sides of the debate have evidence to support them, and it is still an open discussion. The Tully Monster isn’t just unclassified. It’s actively resistant to classification, which is a different and far stranger problem.
Fast Facts
- Discovered in 1958 in Illinois’ Mazon Creek formation by amateur collector Francis Tully
- Lived approximately 300 million years ago during the Carboniferous period
- Grew up to roughly 35 cm (about 14 inches) in length
- Over 1,200 specimens have been formally studied – and consensus still hasn’t arrived
- Illinois named it the official State Fossil in 1989, despite no one knowing what it actually is
#14 – Hallucigenia’s Upside-Down Reconstruction Nightmare

Hallucigenia sparsa earned its name honestly. When paleontologist Simon Conway Morris first reconstructed it in 1977, he had it completely upside down – walking on its spines and waving its legs in the air like antennae – and the error held for over a decade before anyone caught it. Even after it was flipped right-side up, things didn’t get much clearer. This Cambrian creature from Canada’s Burgess Shale has seven pairs of rigid spines along its back, seven pairs of legs tipped with claws, and a head that was initially mistaken for a tail because it’s so featureless from certain angles.
Its evolutionary relationships have bounced between velvet worms, arthropods, and a proposed separate phylum depending on which features the researchers weighted most. The claws at the tips of its legs do match those of modern velvet worms with enough precision to suggest a connection, but the rest of the body layout doesn’t follow through. Related species from China’s Chengjiang fossil beds have added detail without adding clarity. What Hallucigenia tells us most clearly is that early animal evolution was far messier and more experimental than any clean family tree suggests.
#13 – Opabinia’s Five-Eyed Proboscis Puzzle

When Harry Whittington presented his reconstruction of Opabinia regalis at a 1972 paleontology symposium, the audience reportedly laughed. Not out of mockery – out of sheer disbelief. Five eyes mounted on stalks. A single flexible proboscis ending in a claw, pointing backward toward the mouth. Paddle-like lobes along the sides of a soft, segmented body. It looked like someone had described an animal to an artist who had never seen any animal. Yet the fossils were unambiguous. This thing was real, it lived, and it apparently thrived in Cambrian seas around 505 million years ago.
The debate over where it belongs has never fully resolved. Early work suggested it might be a primitive arthropod ancestor, which would make it distantly related to insects, crabs, and spiders. But its combination of features – particularly that unique five-eyed arrangement and the backward-facing proboscis – appears nowhere else in the fossil record, before or after. No other animal has ever been found with that exact configuration. Whether Opabinia represents a genuine dead-end experiment or a distant relative of a known group that simply hasn’t been traced yet remains an open argument with no resolution in sight.
#12 – Prototaxites’ Giant Unknown Life Form

Picture a Devonian landscape roughly 420 million years ago: low-growing plants, early insects, and rising from the ground like ancient smokestacks, enormous column-shaped organisms reaching up to 26 feet tall. These were Prototaxites, and for most of the time since their discovery in the 1800s, everyone assumed they were some kind of giant tree or conifer. Then researchers looked more carefully and realized there were no rings, no leaves, no bark – none of the structures that define a plant. The tree hypothesis collapsed.
The giant fungus hypothesis replaced it for a while, and it held enough ground that some textbooks still carry it. But chemical and cellular analysis has since complicated even that interpretation. The internal structure shows a multicellular organization that doesn’t cleanly match fungal tissue, and the carbon isotope ratios from different specimens vary in ways that don’t fit a single organism type. One serious proposal is that Prototaxites wasn’t a single kind of organism at all, but a composite – something like a massive lichen, a partnership between unrelated life forms. If that’s true, it may represent an entirely extinct ecological strategy with no living parallel.
At a Glance
- Up to 26 feet tall – the largest known land organism of the Devonian period
- Existed for roughly 40 million years before vanishing from the fossil record
- No roots, leaves, bark, or reproductive structures have been identified
- Carbon isotope signatures vary so much between specimens that some researchers suspect it wasn’t one type of organism at all
- Has been formally classified as a tree, a fungus, a lichen, and an alga – none has stuck
#11 – Dickinsonia’s Ediacaran Placement Problem
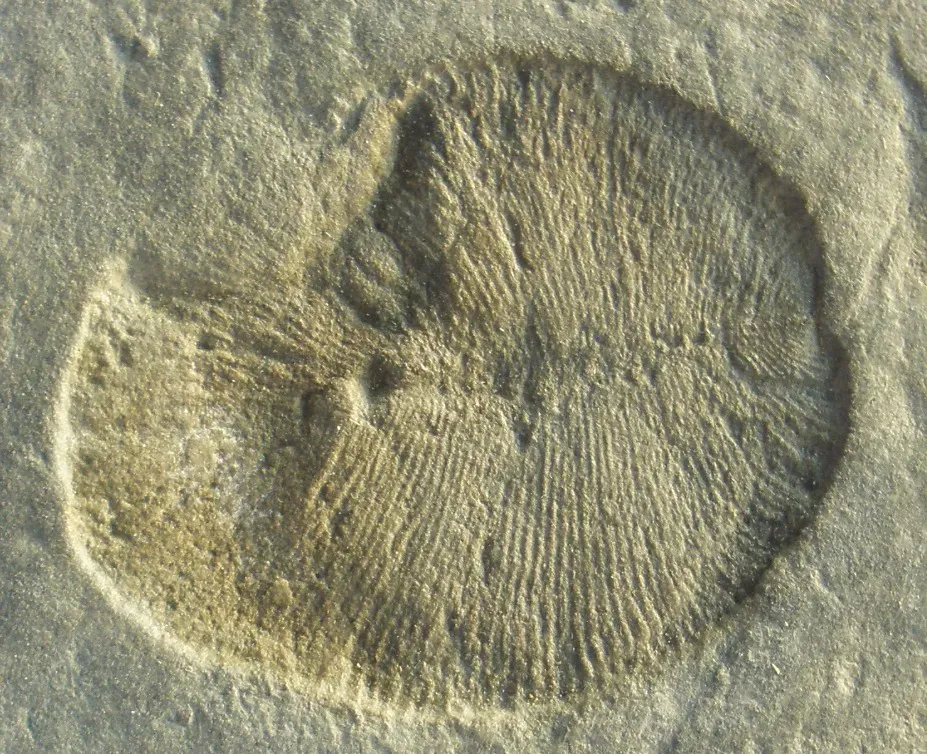
Dickinsonia fossils look, at first glance, like flattened oval rugs. Ribbed, bilaterally symmetrical, sometimes more than a meter long, they appear in 550-million-year-old rocks from Australia and Russia in remarkable numbers. For years the dominant assumption was that they were some kind of primitive animal – maybe a relative of flatworms or early cnidarians. Then trace fossils appeared suggesting Dickinsonia could move, which pointed toward animal behavior. A 2018 study detected cholesterol molecules preserved in one specimen, which seemed to confirm animal biochemistry and briefly made headlines worldwide.
But the picture is still not clean. Dickinsonia’s growth pattern – the way new segments appeared at one end while existing ones shifted – matches no living animal group and no known developmental program in biology. Some researchers have seriously proposed that Dickinsonia and its Ediacaran relatives weren’t animals, fungi, or plants, but members of an entirely separate kingdom of multicellular life that rose to global dominance and then went extinct without leaving descendants. If that interpretation holds up, then for tens of millions of years, Earth was dominated by complex life that has no relatives alive today at all.
#10 – Charnia’s Rangeomorph Affinities

Charnia masoni holds an important title in the history of paleontology: the first fossil from Precambrian rocks to be accepted by mainstream science as a genuine organism rather than a mineral artifact. A 15-year-old schoolgirl named Tina Negus spotted it in Charnwood Forest, England, in 1957, but her discovery was initially dismissed by a geologist who insisted complex life couldn’t exist that far back. When Roger Mason found another specimen the same year and published it, the scientific community had to accept that complex multicellular life existed 570 million years ago – pushing the origin of life’s complexity dramatically further back than anyone had believed.
What Charnia actually was, though, remains stubbornly unclear. It looks like a frond or sea pen, with fractal branching that repeats the same pattern at every scale like a natural fern. But unlike sea pens, there’s no evidence of a mouth, a gut, a feeding current, or any way to capture food. The current leading theory is that it absorbed dissolved organic nutrients directly from the surrounding water – a feeding strategy called osmotrophy – which, if true, means it operated on entirely different biological principles than almost any living complex organism today.
#9 – Anomalocaris and Its Disassembled Identity

Anomalocaris – “abnormal shrimp” in Latin – got its name from a mistake. Its grasping frontal appendages were discovered first and identified as the tail of a shrimp. Its circular mouth was found separately and described as a jellyfish. Its body was classified as yet another creature entirely. It took decades of finds before someone realized all these “different animals” were parts of the same predator, one that reached three feet long and prowled Cambrian seas as probably the apex predator of its time. It was a humbling lesson in how wrong fragmentary evidence can lead you.
Even after the reassembly, Anomalocaris keeps generating new arguments. It’s generally placed in a group called radiodonts, loosely related to modern arthropods, but exactly where it slots into the arthropod family tree is still debated. Its compound eyes are among the largest and most acute to have ever existed – each eye is up to 3 centimetres long and contains more than 16,000 lenses. The existence of highly visual hunters during the Cambrian would have accelerated the pace of the predator–prey arms race then under way. Its grasping appendages show sophisticated multi-joint articulation that wouldn’t reappear in the record for hundreds of millions of years. The more specimens turn up, the more impressive it becomes, and the less certain its exact relationships remain.
Why It Stands Out
- Each compound eye packed over 16,000 individual lenses – rivalling the visual acuity of modern dragonflies
- Fossils push the origin of compound eyes deeper down the arthropod stem lineage and indicate the compound eye evolved before hardened exoskeletons
- Body parts were initially described as three completely separate animals before being correctly unified
- Reached up to about 1 meter in length – the largest known predator of the Cambrian seas
- Its multi-jointed grasping appendages represent engineering complexity with no known earlier precedent
#8 – Marrella and the Dead End of the “Lace Crab”

Marrella splendens is actually one of the most abundant animals in the Burgess Shale – it makes up roughly 70% of all specimens from that deposit. Charles Doolittle Walcott, who discovered the shale in 1909, initially called it a “lace crab” and filed it loosely with trilobites. It’s a delicate, beautiful thing under a lens: feathery gills, two pairs of long sweeping head spines, and fine appendages that give it an almost ornate appearance. You’d expect that something so abundant and so well-preserved would be easy to classify.
It isn’t. Detailed work by Harry Whittington in the 1970s revealed that Marrella doesn’t fit with trilobites, crustaceans, or any other established arthropod group. It sits in its own isolated position as a primitive arthropod outside all the major living branches – a lineage that flourished in Cambrian seas and then simply stopped. No descendants. No later relatives. One of the most common animals of its time left no traceable legacy. Marrella is a reminder that evolutionary success in one era is no guarantee of survival into the next, and that the Cambrian was full of experiments that the world eventually stopped running.
#7 – Wiwaxia’s Scale Armor Mystery

Wiwaxia corrugata looked, in life, like a small oval pincushion. It moved slowly across the Cambrian seafloor covered in overlapping mineralized scales and armed with two rows of long defensive spines rising from its back. The armor is striking and clearly functional, but when paleontologists tried to figure out what kind of animal it was protecting, the arguments started immediately and haven’t stopped. The scale structure draws comparisons to mollusks. The spines look more like those of certain annelid worms. Its feeding apparatus – a rasping tongue-like structure called a radula – closely resembles that of modern mollusks.
The mismatch between those features is the core of the problem. If Wiwaxia is a mollusk, its armor is wrong. If it’s an annelid, its feeding apparatus is wrong. Some researchers have proposed it belongs in its own separate lineage that was ancestral to both groups – a “stem lophotrochozoan” that predated the split between mollusks and worms. Others argue that’s overreaching and that it belongs firmly in one camp or the other. The specimen quality from the Burgess Shale is exceptional, which makes the disagreement all the more striking. This isn’t a case of bad preservation. The animal is just genuinely strange.
Quick Compare
- Scale structure: Points toward mollusks
- Defensive spines: Resemble annelid worm features
- Feeding apparatus (radula): Closely matches modern mollusks
- Overall body plan: Matches neither group cleanly
- Current best guess: A “stem lophotrochozoan” – ancestral to both, belonging to neither
#6 – Nanotyrannus and the Tyrannosaur Identity War

Few debates in paleontology generate as much heat as the Nanotyrannus question. The argument is simple on its surface: are the slender-skulled tyrannosaur fossils from the Hell Creek Formation a separate species of small, fast predator, or are they just juvenile T. rex specimens that haven’t finished growing yet? The name Nanotyrannus lancensis was proposed in 1988, and the debate raged for decades with no sign of the two camps reaching a truce. At stake wasn’t just taxonomy – it was the question of whether North America’s late Cretaceous had two large tyrannosaur species sharing the same ecosystem.
That debate has now taken a decisive turn. Paleontologists confirmed in a landmark study published in October 2025 in Nature that Nanotyrannus is a distinct dinosaur genus – not merely a juvenile Tyrannosaurus rex – providing compelling evidence that it was a separate species that coexisted with T. rex during the late Cretaceous period, approximately 66 million years ago. Using growth rings, spinal fusion data, and developmental anatomy, the researchers demonstrated that the specimen was around 20 years old and physically mature – and that its skeletal features, including larger forelimbs, more teeth, fewer tail vertebrae, and distinct skull nerve patterns, are biologically incompatible with Tyrannosaurus rex. Confirmation of Nanotyrannus means that predator diversity in the last million years of the Cretaceous was much higher than previously thought, and hints that other small-bodied dinosaur species might also be victims of mistaken identity.
#5 – Spriggina’s Place at the Edge of the Animal Kingdom

Spriggina floundersi, named after South Australian geologist Reg Sprigg who discovered it in the 1940s, is one of the oldest fossils ever suggested as a possible animal ancestor. At 550 million years old, it shows something remarkable: clear, repeated body segments and a distinct crescent-shaped head shield that looks almost arthropod-like. In a world of vague Ediacaran blob-shapes, Spriggina stands out for having something that at least resembles organized animal architecture. It’s been proposed as an ancestor of trilobites, as an early annelid relative, and as a bilaterian animal in the broadest sense.
The problem is that the more carefully researchers examine it, the less certain each of those links becomes. There are no preserved appendages, no gut trace, no clear mouth structure. The head shield is unusual enough that some paleontologists argue it has no real equivalent in any later animal group, which would mean Spriggina isn’t an ancestor of anything we know – it’s another Ediacaran experiment that ended without continuation. Whether it sits just inside or just outside the animal kingdom’s origin point is a question with enormous implications for understanding when and how animals first evolved, and right now the answer is genuinely uncertain.
#4 – The “Godzillus” Cincinnati Mystery

In 2011, amateur fossil hunter Ron Fine pulled something out of Ordovician-age rocks near Cincinnati, Ohio, that stopped professional paleontologists cold. The specimen – eventually nicknamed “Godzillus” by its finders – is roughly six feet long, heavily lobed and irregular in shape, and dates to approximately 450 million years ago. Fine presented it at a regional paleontological conference expecting someone to immediately tell him what it was. Nobody could. Experts examined it and ruled out trilobites, mollusks, algae mats, and coral colonies. It didn’t match anything in the Ordovician fossil record.
The formal scientific study of the specimen has proceeded cautiously, partly because the preservation is unusual and partly because the organism’s structure doesn’t lend itself to easy tissue analysis. What’s clear is that it represents a large, complex organism with a body plan that left no obvious relatives in later rocks. The Ordovician was itself a period of explosive diversification – the “Great Ordovician Biodiversification Event” – so the existence of unknown body plans from that era isn’t shocking in theory. But having a six-foot physical specimen sitting in front of you that nobody can name is a different kind of confrontation with uncertainty than a theoretical gap in a phylogeny chart.
#3 – Pikaia and the Shaky Foundation of Our Own Phylum

Pikaia gracilens used to be a feel-good story in paleontology. Discovered in the Burgess Shale and described formally in the 1970s, this small, ribbon-like Cambrian swimmer appeared to show a notochord – the stiffening rod that defines the chordate phylum, the group that includes vertebrates and, ultimately, humans. For years it was celebrated as the earliest known member of our lineage, a 505-million-year-old ancestor swimming in the same Cambrian seas as Opabinia and Anomalocaris. Stephen Jay Gould famously used it as an example in Wonderful Life, musing that if Pikaia had gone extinct, humans might never have existed.
Replay the tape of life and the chance becomes vanishingly small that anything like human intelligence would grace the replay.
Stephen Jay Gould, Wonderful Life
The problem is that Pikaia’s chordate credentials have been steadily eroding. Detailed reanalysis of its musculature and internal anatomy suggests that what was interpreted as a notochord may actually be a different kind of internal support structure – one that doesn’t qualify for chordate membership. More confidently identified early chordates have since been found in Chinese Cambrian deposits, pushing Pikaia to the margins of the story. It may still be an early chordate. It may sit just outside the phylum entirely. Either way, the animal we once thought anchored our deepest origins is now a question mark, which has a way of making the ground feel less stable than it did before.
#2 – Rangeomorphs and the Architecture That Disappeared Forever

Rangeomorphs were everywhere during the Ediacaran period, carpeting deep-sea floors with fractal fronds that repeated the same branching geometry at every scale, from the full organism down to the tiniest visible filament. Some grew to nearly two meters. They built entire communities, lived apparently without mouths, guts, or any recognizable organ system, and then – right at the boundary between the Ediacaran and Cambrian periods, around 541 million years ago – every single species vanished. Not a reduction. Not a gradual decline. Complete disappearance, with no descendants in the Cambrian or any later period.
Their proposed feeding strategy is osmotrophy: absorbing dissolved organic molecules directly through the body surface, which would have worked in an Ediacaran ocean rich in that kind of chemistry but may have become nonviable as ocean conditions changed at the Cambrian boundary. What makes rangeomorphs particularly haunting is that their fractal growth architecture has never reappeared in the fossil record. No later organism – animal, plant, or fungus – builds itself the same way. They represent a biological design principle that existed, succeeded on a global scale, and was erased so completely that evolution never returned to it. That’s not just an extinction. That’s an entire way of being alive that the world tried once and never tried again.
Worth Knowing
- Some rangeomorphs grew to nearly 2 meters – large and structurally complex, yet with no mouth, gut, or organs
- Their fractal branching geometry repeats identically from the whole organism down to the finest filament – a design found nowhere else in nature
- They vanished precisely at the Ediacaran–Cambrian boundary (~541 million years ago), with zero known descendants
- Osmotrophy – absorbing nutrients directly through body surfaces – is their best-supported feeding strategy, and it may have become unviable as ocean chemistry shifted
- No animal, plant, or fungus alive today builds itself using the same architectural principle
#1 – The Cambrian Explosion’s Unresolved Trigger

Everything else on this list flows from this one. Roughly 538 million years ago, over a geologically brief window of perhaps 20 to 25 million years, the fossil record goes from sparse and strange to suddenly packed with complex animals bearing eyes, limbs, shells, guts, and nervous systems. Arthropods, mollusks, echinoderms, chordates – most of the major animal body plans that exist today appeared in this single interval without clear Precambrian precursors in the hard fossil record. Biologists call it the Cambrian Explosion, and despite 150 years of focused study, no one can fully explain why it happened when it did or as fast as it did.
The proposed triggers form a long and still-contested list. A 2024 study in Nature Geoscience from an international consortium of scientists from more than 50 institutions proposed that the Cambrian Explosion may have been triggered by only a small increase in oxygen levels in Earth’s atmosphere and shallow ocean waters. “It’s not a huge increase in oxygen, but it might be enough to cross critical ecological thresholds, based on what we see in modern areas with naturally low oxygen,” noted Stanford researcher Erik Sperling. Yet even that theory has taken hits: a separate paper in the Proceedings of the National Academy of Sciences suggests oxygen might have been quite high long before the Cambrian period, casting doubt on the idea that it played a role as a trigger. Making the problem sharper, molecular clock studies – which estimate divergence times from genetic mutation rates in living animals – consistently push the origin of major animal groups back well before the Cambrian, into the Ediacaran or earlier, while the fossil record shows almost nothing complex there. Either the fossils are lying, the molecular clocks are wrong, or the Precambrian ancestors existed as soft, small, or rare organisms that simply didn’t preserve. All three remain live possibilities. The Cambrian Explosion isn’t just one mystery. It’s the mystery that every other mystery on this list feeds into.
Quick Compare: Leading Trigger Theories
- Oxygen threshold rise: Supported by ocean sediment data; challenged by evidence of high pre-Cambrian oxygen levels
- Predator–prey arms race: First predators drove defensive evolution across all prey lineages – a compelling feedback loop
- Snowball Earth thaw: Melting glaciers flooded oceans with nutrients, potentially fueling a burst of diversification
- Great Unconformity: A global erosion event may have released continental minerals that triggered biomineralization
- Genetic regulatory innovation: New “toolkit” genes may have unlocked entirely new body plan possibilities simultaneously
The Bigger Picture

What these 15 fossils share isn’t just strangeness. It’s the specific kind of strangeness that comes from life experimenting before it settled into the patterns we now take for granted. The Cambrian and Ediacaran weren’t just early chapters in a story that was always heading toward us – they were periods when the rules of life were still being written, when body plans that seem impossible today were not only possible but thriving.
The honest opinion here is that some of these mysteries will stay mysteries. The Tully Monster may never get a final answer – current findings strongly indicate it was an invertebrate, though its exact classification remains unresolved. Rangeomorphs may be too alien to ever fully map onto living biology. And that’s not a failure of science – it’s a feature of deep time. The further back you go, the less the present can tell you about what the past was doing. The fossils that resist explanation aren’t gaps in our knowledge waiting to be filled. They’re evidence that life has been stranger, more inventive, and more ruthlessly experimental than anything we currently have the vocabulary to describe. The fact that we keep digging anyway is the most human thing about all of this.

